Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Антиоксидантная защита мозга

2
Особенности окислительного метаболизма мозга Высокий уровень потребления кислорода Большое количество липидов с ненасыщеными жирнокислотными радикалами Насыщенность железом белков- переносчиков Низкий уровень антиоксидантной защиты

3
БАЛАНС АФК В ЖИВЫХ КЛЕТКАХ ТУШЕНИЕ АФК ТУШЕНИЕ АФК ГЕНЕРАЦИЯ АФК ГЕНЕРАЦИЯ АФК Дыхательная цепь митохондрий, NADPH-оксидаза нейтрофилов, микросомальное окисление, неферментативное окисление биогенных аминов СОД, Каталаза, Пероксидазы, Низкомолек. антиоксиданты (мочевая кислота, таурин, витамины A, C, E, карнозин, N- ацетилцистеин, глутатион), xелаторы ионов железа СОД, Каталаза, Пероксидазы, Низкомолек. антиоксиданты (мочевая кислота, таурин, витамины A, C, E, карнозин, N- ацетилцистеин, глутатион), xелаторы ионов железа

4
Нейроденеративные процессы Старение АФК AO система AO система ДЕФИЦИТ антиокси- дантов Метаболические нарушения Токсиканты Тяжелые металлы ОКИСЛИТЕЛЬНЫЙ С Т Р Е С С

6
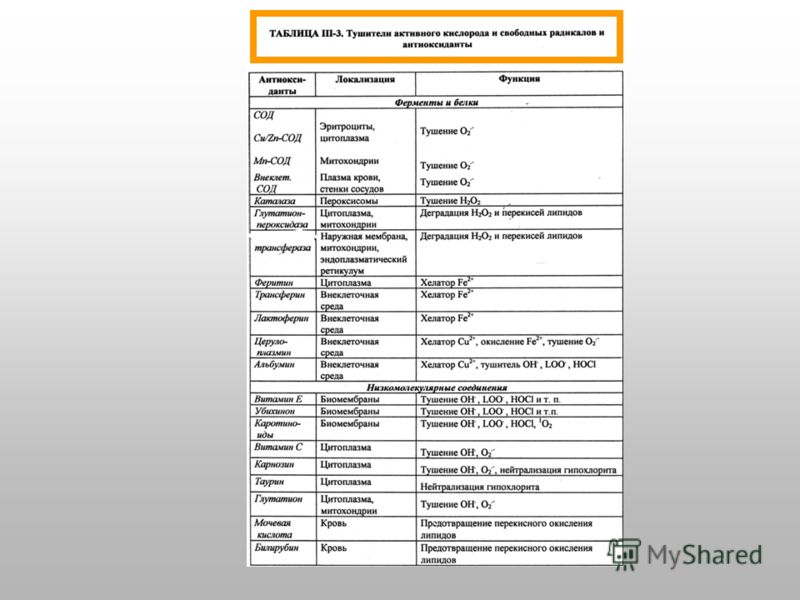
Антиоксидантные ферменты и низкомолекулярные антиоксиданты Супероксиддисмутаза (разные формы содержат Cu/Zn и Mn ): О О Н + Н 2 О 2 + О 2 Каталаза ( гемосодержащий фермент ): 2Н 2 О 2 2Н 2 О + О 2 Глутатионпероксидаза ( содержит остаток селеноцистеина ): 2GSH + Н 2 О 2 GSSG + 2Н 2 O Глутатионредуктаза (содержит FAD): GSSG + 2НАДФН 2GSH + 2НАДФ Токоферол (вит. Е) Ретинол (вит. А) Аскорбиновая кислота (вит. С) Глутатион восстановленный N-ацетилцистеин Мочевина, мочевая кислота Карнозин и другие гистидинсодержащие дипептиды
: О 2 - + О 2 - + 2Н + Н 2 О 2 + О 2 Каталаза ( гемосодержащий фермент ): 2Н 2 О 2 2Н 2 О + О 2 Глутатионпероксидаза ( содержит остаток")
7
РЕГУЛЯЦИЯ

8
Убиквити- нилирование и деградация поврежденных молекул белка

9
Контроль уровня АФК клетками глии Соотношение глия/нейроны растет в онтогенезе от 0,2 до 1,6 (человек) Соотношение глия/нейрон в мозге Эйнштейна составляло 1,95
 Соотношение глия/нейрон в мозге Эйнштейна составляло 1,95")
10
Роль каталазы Н202Н202 Н202Н202 Время Контрольная культураKnock-out глиальных клетокGlu-peroxidase -/- + BSO (ингибитор глу-пероксидазы) +/- 3-АТ (ингибитор каталазы) + 3-АТ +/- ВSО
 +/- 3-АТ (ингибитор каталазы) + 3-АТ +/- ВSО")
11
GSH -L-glutamyl-L-cysteinylglycine В клетках млекопитающих концентрация от мМ В мозге ~ мМ, в межклеточном пространстве присутствует в микромолярной концентрации Не проникает через гематоэнцефалический барьер (вопрос о специфическом переносчике открыт) Синтезируется из проникающих в клетку предшественников – глутамата, цистеина и глицина Метаболизм GSH имеет тонкие различия в клетках мозга разного типа (астроглия поддерживает необходимый уровень предшественников для синтеза GSH в нейронах) В синтезе принимают участие ферменты – γGluCys синтетаза и глутатион синтетаза Конечный продукт окисления – глутатион дисульфид (GSSG), восстанавливается глутатионредуктазой (NADPH), GSH/GSSG порядка 1000/1 Уровень внутриклеточного глутатиона изменяется при патологиях (показано снижение уровня на 40-50% при болезни Паркинсона и, наоборот, возрастание при гомоцистеинемии)

12
Функции GSH в клетках Антиоксидантная - прямое взаимодействие с радикалами в неэнзиматических реакциях (Saer et al.,1990; Winterbourn, 1994); донор электронов в реакциях восстановления перекисей, катализируемых глутатион пероксидазами (Chance et al., 1979) Обеспечивает поддержание тиолового статуса клетки путем сохранения сульфгидрильных групп в восстановленном состоянии (Cotdrave and Gudes, 1997) Участник процесса детоксикации ксенобиотиков, кофактор в реакциях изомеризации, форма хранения и транспортировки цистеина (Meister and Anderson, 1983; Cooper, 1997) Участник процессов пролиферации (Pool et al,. 1995) Участие в регуляции апоптоза (Chibelli et al., 1998; Hall, 1999) NEW! Является нейротрансмиттером и нейромодулятором (в микромолярных концентрациях является агонистом глутаматных рецепторов; в миллимолярных концентрациях модулирует SH – группы NMDA рецепторов) (Janaky et al., 1999) NEW! При определенных условиях может выступать в качестве прооксиданта (Paolicchi et al., 2002)
; донор электронов в реакциях восстановления перекисей, катализируемых глутатион пероксидазами (Chance et al.,")
13
Способы изменения содержания глутатиона в клетках in vitro - GSH CDNB (chloro-2,4- dinitrobenzene) цитозоль+ митохондрии DEM (diethyl maleimide) цитозоль образуют конъюгаты с GSH в результате реакции, катализируемой глутатион-S- трансферазой +GSH использовали et-GSH (легко проникает в клетку благодаря этерифицированной группе глицинового остатка и деэтерифицируется внутриклеточно) DEM –1 mM, 1h CDNB - 1 mM, 30 min Et-GSH – 5 mM, 30 min N=5 Уровень GSH оценивали цитометрически (непосредственно в живых клетках) – с помощью флуоресцентной краски на глутатион – CMFDА (chloromethyl fluorescein )
 цитозоль+ митохондрии DEM (diethyl maleimide) цитозоль образуют конъюгаты с GSH в результате реакции, катализируемой глутатион-S- трансферазой +GSH исп")
14
Истощение цитозольного и митохондриального пулов GSH при помощи CDNB приводит к увеличению генерации митохондриальных АФК, снижает уровень АТФ в клетке, снижает транспортную активность Na,K-АТФазы, и, в конечном итоге, резко понижает жизнеспособность клеток + CDNB 123(DHR) – dihydrorhodamine, окисляется до катиона rhodamine 123, накапливается в митохондриях АФК АТФ

15
Этанол

16
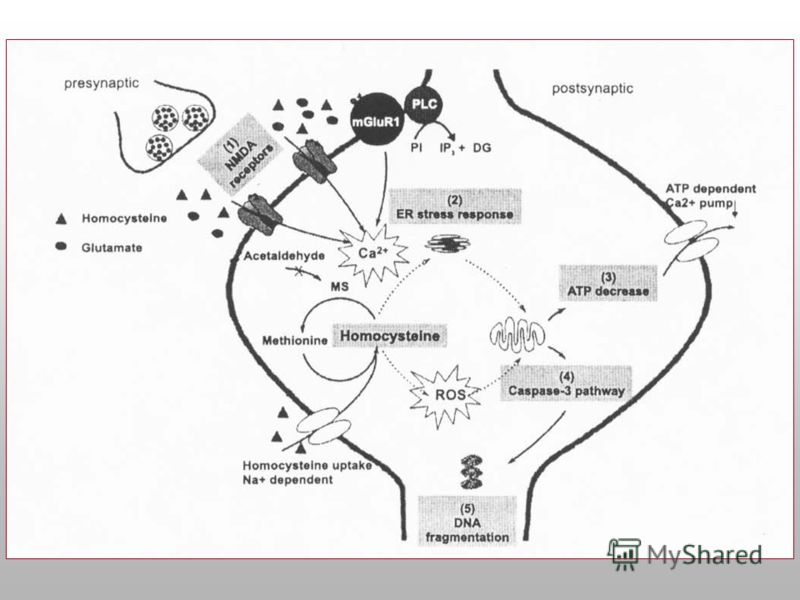
Гомоцистеин (ГЦ) представляет собой серосодержащую аминокислоту история исследований, связанных с определением гомоцистеина, начинается с 1932 г., когда De Vigneaud обнаружил эту аминокислоту как продукт деметилирования метионина в организме активно участвует в окислительно-восстановительных реакциях, он способен к аутоокислению, в результате которого образуется гомоцистеиновая кислота [Welch G., 1998]. вне клетки находится либо в окисленной форме (1%), либо в связанном с белками состоянии (70%). в понятие «общий гомоцистеин» входят все формы гомоцистеина, циркулирующие в плазме крови [Шевченко О.П., Олефриенко Г.А., 2002]. диагноз гипергомоцистеинемии ставят в том случае, если уровень гомоцистеина в крови превышает 15 мкмоль/л. Концентрация гомоцистеина в плазме крови в пределах 15–30 мкмоль/л свидетельствует об умеренной гипергомоцистеинемии, от 30 до 100 мкмоль/л – о промежуточной, 100 – 500 мкмоль/л – тяжелой [Welch G., Loscalo J., 1998; Warren C., 2002].
 представляет собой серосодержащую аминокислоту история исследований, связанных с определением гомоцистеина, начинается с 1932 г., когда De Vigneaud обнаружил эту аминокислоту как продукт деметилирования метионина в организме активно")
17
ГОМОЦИСТЕИН КАК ФАКТОР РИСКА ОКИСЛИТЕЛЬНОГО СТРЕССА МОЗГА И СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЫ ГОМОЦИСТЕИН ГОМОЦИСТЕИНОВАЯ КИСЛОТА

18
Причины и следствия повышения уровня гомоцистеина в плазме крови Развитие седечно- сосудистых патологи й Тромбообразование (риск тромбоэмболии увеличивается в несколько раз) Атеросклероз в 1975 г. McCully предложил гомоцистеиновую теорию атеросклероза Нейро-дегенеративные заболевания (болезнь Альцгеймера) Нарушение течения беременности и формирования плода (главное - патологии развития нервной системы) Нарушение какого - либо из этапов превращения ГЦ (вследствие недостатка витаминов или генетического дефекта ферментов) Сопутствующие заболевания (почечная недостаточность) Действие приема некоторых лекарственных препаратов
 Атеросклероз в 1975 г. McCully предложил гомоцистеиновую теорию атеросклероз")
20
Молекулярные последствия гипергомоцистеинемии Интенсификация метилирования нуклеиновых кислот, белков и фосфолипидов Повышенный внутриклеточный уровень свободных радикалов Модификация глутаматных рецепторов

21
In vitro

22
PI (пропидий иодид ) ex =485 нм, em =610нм (оценка смертности) DCFH-DA (2,7 – дихлордигидрофлуоресцеин) exc =485 нм, em =530нм (оценка АФК) Fluo–3 АМ exc =488 нм, em =530нм (оценка цитоплазматического кальция) Аннексин V – FITC exc =488 нм, em =530нм (оценка степени экспонирования фосфатидилсерина на начальных стадиях апоптоза) В работе использовались следующие флуоресцентные зонды: Исследовали действие ГЦ и ГЦК на глутаматные рецепторы нейронов и лимфоцитов in vitro методом проточной цитометрии
 ex =485 нм, em =610нм (оценка смертности) DCFH-DA (2,7 – дихлордигидрофлуоресцеин) exc =485 нм, em =530нм (оценка АФК) Fluo–3 АМ exc =488 нм, em =530нм (оценка цитоплазматического кальция) Аннексин V – FITC exc =488 нм, em =530нм")
23
Действие ГЦ и ГЦК на глутаматные рецепторы нейронов in vitro КальцийАФК

24
ГЦ и ГЦК способны взаимодействовать как с ионотропными, так и с метаботропными глутаматными рецепторами AIDA – антагонист метаботропных рецепторов I класса MSOP – антагонист метаботропных рецепторов III класса MK-801 – антагонист ионотропных рецепторов

25
In vivo

26
Experimental protocol ± Treatment with possible protector Pregnant rat Pups Methionine with drinking water 1 g/kg dayly 2 weeks 4 weeks Cytometric test Behavioral test 1818

27
Модель пренатальной гипергомоцистеинемии ГруппаДиетаКоличество семей Среднее количество особей в помете Вес (г) (для потомства в возрасте 10 дней) Группа 1 (контрольные) обычная диета 6 14 ± 2 23,3 ± 0,4 Группа 2 (метиониновые) введение в питье беременных животных метионина (из расчета 1±0,01 г/кг веса с учетом объема потребляемой жидкости) и ограничение в рационе витаминов группы B и фолиевой кислоты 4 (6 – 2) 7 ± 118,9 ± 0,5 Группа 3 (метионин + карнозин) то же + карнозин из расчета 0,1±0,01 г/кг веса 6 11 ± 222,9 ± 0,9
 (для потомства в возрасте 10 дней) Группа 1 (контрольные) обычная диета 6 14 ± 2 23,3 ± 0,4 Группа 2 (метиониновые) введение в питье берем")
28
Content of HC in the blood of rats under methionine over-loading Control8-13 mkM Methionine overload48-52 mkM

29
Определение чувствительности глутаматных рецепторов к лигандам 1.У животных, получавших метионин, наблюдается тенденция к снижению чувствительности глутаматных рецепторов. 2.Рецепторы «метиониновых» животных утратили чувствительность к NMDA, однако чувствительность к HC и HCA сохранилась. Нейроны преинкубировали с NMDA, HCA и HC в концентрации 500 мкМ 30 мин

30
В случае активации нейронов при инкубации с HC или HCA ответ нейронов реализуется через не-NMDA глутаматные рецепторы AIDA - антагонист mGlu 1 MSOP – антагонист mGlu3

31
При обработке данных использовали специальную программу, которая позволяет оценить следующие параметры: 1) время от начала движения крысы в бассейне до достижения ею платформы (в сек); 2) длину пути (в метрах); 3) среднюю скорость (в м/c); 4) сколько времени крыса плавала с быстрой, средней и медленной скоростью (в % от всего времени прохождения теста); 5) время нахождения крысы в центре бассейна (внутренний круг) или около бортика (внешний круг), что также позволяет оценить характер поисков Тест Морриса R.G. M. Morris et al Nature, 297, Проводили для животных из каждой семьи в возрасте месяцев. Животные содержались на диете, соответствующей каждой группе.
 время от начала движения крысы в бассейне до достижения ею платформы (в сек); 2) длину пути (в метрах); 3) среднюю скорость (в м/c); 4) сколько")
32
Анализ результата теста Морриса Регистрируемый параметр (данные представлены для четвертой попытки) Группа 1 (контрольные) Группа 2 (метиониновые) Группа 3 (карнозиновые) Время нахождения платформы, с 20 ± 7140 ± 1845 ± 6 Длина траектории, м 5 ± 220 ± 58 ± 2 Средняя скорость, м/c 0,24 ± 0,020,18 ± 0 020,25 ± 0,04 Время нахождения в центральной области бассейна, в % от всего времени 20 ± 77 ± 535 ± 5 Оценивали пространственную ориентацию животных: в первый день эксперимента осуществляется претренинг животных во второй день животным предоставляется 4 -5 попыток найти платформу
 Группа 1 (контрольные) Группа 2 (метиониновые) Группа 3 (карнозиновые) Время нахождения платформы, с 20 ± 7140 ± 1845 ± 6 Длина траектории, м 5 ± 220")
33
Антиоксидантные ферменты и низкомолекулярные антиоксиданты Супероксиддисмутаза (разные формы содержат Cu/Zn и Mn ): О О Н + Н 2 О 2 + О 2 Каталаза ( гемосодержащий фермент ): 2Н 2 О 2 2Н 2 О + О 2 Глутатионпероксидаза ( содержит остаток селеноцистеина ): 2GSH + Н 2 О 2 GSSG + 2Н 2 O Глутатионредуктаза (содержит FAD): GSSG + 2НАДФН 2GSH + 2НАДФ Токоферол (вит. Е) Ретинол (вит. А) Аскорбиновая кислота (вит. С) Глутатион восстановленный N-ацетилцистеин Мочевина, мочевая кислота Карнозин и другие гистидинсодержащие дипептиды
: О 2 - + О 2 - + 2Н + Н 2 О 2 + О 2 Каталаза ( гемосодержащий фермент ): 2Н 2 О 2 2Н 2 О + О 2 Глутатионпероксидаза ( содержит остаток")
40
Парадоксальное увеличение генерации АФК в нейронах в ответ на увеличение содержания GSH может быть связано с прооксидантным действием GSH 1) GSH+ Cu 2+ /Fe 3+ Cu + /Fe 2+ + GS· + H + (2) Cu + /Fe 2+ + O 2 Cu 2+ /Fe 3+ + O 2 ·- (3) GS · + GSH GSSG ·- + H + (4) GSSG·- + O 2 O 2 ·- + GSSG (5) 2O 2 ·- + 2H + H 2 O 2 + O 2 (6) H 2 O 2 + Cu + /Fe 2+ OH ·- + OH -
 GSH+ Cu 2+ /Fe 3+ Cu + /Fe 2+ + GS· + H + (2) Cu + /Fe 2+ + O 2 Cu 2+ /Fe 3+ + O 2 ·- (3) GS · + GSH GSSG ·-")
41
Развитие окислительного стресса в нейроне. Регистрация АФК методом проточной цитометрии О 2 е- О 2 - е- Н 2 О 2 е- ОН - е- + 2Н+ Н 2 О + 2Н + Н 2 О 2 + О 2 О 2 + ОН - + ОН (реакция Хабера-Вайса) + О 2 - ОН - + ОН - + Fe 3+ (реакция Фентон) + NO ONOO - + Fe 2+ DCFH 2 DCF Не флуоресцирую- щая молекула DCF проникает в клетку, а ее окисленная форма флуоресцирует
 + О 2 - ОН - + ОН - + Fe 3+ (реакция Фентон) + NO ONOO")
42
Влияние 100 µM уабаина на внутрикеточный уровень АФК в грануляных клетках мозжечка ROS signal PI versus DCF coordinates PI DCF Boldyrev et al, Ann NY Acad. Sci, 2003, 814,

43
Изменение содержания АФК в нейронах в условиях активации ионотропных NMDA- рецепторов и метаботропных рецепторов I(3- HPG) и III(ACPD) классов
 и III(ACPD) классов")
44
Effect of Glutamate Ligands on Na/K-ATPase 3-HPG NMDA ACPD

45
D Aktivated PkB, % Нейробластома SH-SY5Y Р АПОПТОЗ Выход цит С из митохондрий в цитоплазму + - Kulikov et al, Biochemistry, submitted АФК (2+3) 1
 1")
46
Na-насос в нейронах регулирует активность МАР киназы Контроль Уабаин Уабаин+D-AP5

47
При инкубации нейронов с уабаином рост МАР киназы зависит от активности NMDA-рецепторов и ионов кальция

48
Активация нейрональной MAPK уабаином требует активного состояния NMDA- рецепторов и реализуется при участии ионов Са и АФК (* соответствует p

49
Корреляция между ингибированием Na/K-АТФазы и активацией МАРК уабаином

50
Участие Na/K-ATФазы в регуляции апоптоза АФК ПkC MAP Kиназа Na + K + Ca Na + K + 1 NMDA IP 3 K ЭПР Ca 2+ NADPHоксидаза - ЭНДОУАБАИН Са 2+ АПОПТОЗ Bcl2 цит с + ПkВПkВ ПkВПkВ - p53P

51
Проблемы антиоксидантной зашиты ишемического мозга Н202 Н202 Fe 2+ OH. Н20 Н20 Glu-SH NAD NADH 2 GSSG Каталаза Пероксидаза Глу-редуктаза СОД

52
Проблемы антиоксидантной зашиты ишемического мозга Н202 Н202 Fe 2+ OH. Н20 Н20 Glu-SH NAD NADH 2 GSSG Каталаза Пероксидаза Глу-редуктаза СОД

53
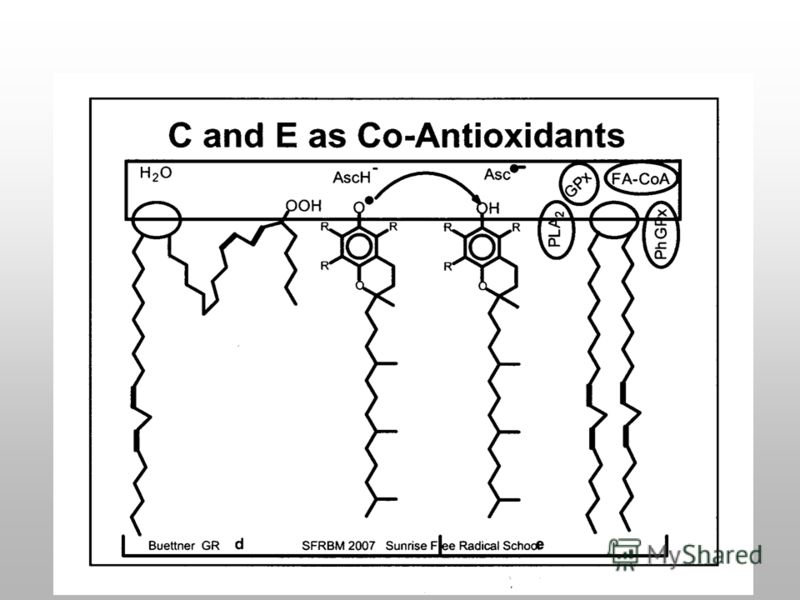
Избыток антиоксидантов вызывает прооксидантный эффект GluSH GS-SG -Tocoferol -Tocoferyl* Ascorbat Asc* АО АО* е*е* АФК NADH 2 NAD

Еще похожие презентации в нашем архиве:













 Натрий (Na) участвует в образовании желудочного сока. Р егулирует выделение почками многих продуктов обмена.")




 - это высокоспецифичные белки, выполняющие функции биологических катализаторов. Катализатор - это вещество, которое ускоряет.")

 в организме человека Работу выполнили студентки 210 группы: Орехова Оксана;")


