Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Лекция 18 ОРГАНЫ ОЩУЩЕНИЯ (продолжение)
")
2
Механорецепторы СЛУХ

3
Ухо External ear –Auricle or pinna: elastic cartilage –External auditory meatus Lined with hairs and ceruminous glands –Tympanic membrane Vibrated by sound waves Middle ear –Auditory or eustachian tube Opens into pharynx, equalizes pressure Ossicles: malleus, incus, stapes: transmit vibrations Oval window

6
Ухо человека

7
External ear: Hearing; terminates at eardrum Middle ear: Hearing; contains auditory ossicles Inner ear: Hearing and balance; interconnecting fluid-filled tunnels and chambers

8
outer ear External ear (a) Middle earInner ear bones of middle ear vestibular system (detects head movement and gravity) auditory nerve to brain auditory canal tympanic membrane oval window (beneath stirrup) round window cochlea auditory tube (Eustachian tube) to pharynx (b) (c)tectoral membrane hair cell basilar membrane axons of auditory nerve hair cells tectorial membrane basilar membrane auditory nerve bony cochlear wall The human ear
 Middle earInner ear bones of middle ear vestibular system (detects head movement and gravity) auditory nerve to brain auditory canal tympanic membrane oval window (beneath stirrup) round window cochlea auditory tube (Eustac")
9
Вестибулярные и слуховые органы человека

10
Scheme (inner ear,crossection):semisircular canal. Hair cells, ampulla,cupula, vestibular nerve, endolymph
:semisircular canal. Hair cells, ampulla,cupula, vestibular nerve, endolymph")
11
Hair cells are the sensory receptors of both the auditory system and the vestibular system in all vertebrates. In mammals, the auditory hair cells are located within the organ of Corti on a thin basilar membrane in the cochlea of the inner ear. They derive their name from the tufts of stereocilia that protrude from the apical surface of the cell, a structure known as the hair bundle, into the scala media, a fluid-filled tube within the cochlea. Mammalian cochlear hair cells come in two anatomically and functionally distinct types: the outer and inner hair cells. Damage to these hair cells results in decreased hearing sensitivity, i.e. sensorineural hearing loss.

12
Механосенсорные волосковые клетки

13
Пучки волосков как детекторы звука и усилители сигнала Внутренние волосковые клетки уха проводят сигнал от звукового колебания до нейронов мозга Внешние волосковые клетки уха - аккустические усилители

14
Пучки волосков как звуковые детекторы и умножители сигнала Outer hair cells do not send neural signals to the brain, but that they mechanically amplify low-level sound that enters the cochlea. The amplification may be powered by movement of their hair bundles, or by an electrically driven motility of their cell bodies. The inner hair cells transform the sound vibrations in the fluids of the cochlea into electrical signals that are then relayed via the auditory nerve to the auditory brainstem and to the auditory cortex. Mammals apparently have conserved an evolutionarily earlier type of hair-cell motility. This so-called hair-bundle motility amplifies sound in all non-mammalian land vertebrates. It is effected by the closing mechanism of the mechanical sensory ion channels at the tips of the hair bundles. Thus, the same hair-bundle mechanism that detects sound vibrations also actively vibrates back and thereby mechanically amplifies weak incoming sound.

15
Внутренние волосковые клетки – преобразование звукового сигнала в нервный импульс The deflection of the hair-cell stereocilia opens mechanically gated ion channels that allow any small, positively charged ions (primarily potassium and calcium) to enter the cell. Unlike many other electrically active cells, the hair cell itself does not fire an action potencial. Instead, the influx of positive ions from the endolymph in Scala media depolarizes the cell, resulting in a receptor potencial. This receptor potential opens voltage gated calcium channels; calcium ions then enter the cell and trigger the release of neurotransmitters at the basal end of the cell. The neurotransmitters diffuse across the narrow space between the hair cell and a nerve terminal, where they then bind to receptors and thus trigger action potentials in the nerve. In this way, the mechanical sound signal is converted into an electrical nerve signal. The repolarization in the hair cell is done in a special manner. The perilymph in Scala tympani has a very low concentration of positive ions. The electrochemical gradient makes the positive ions flow through channels to the perilymph. Hair cells chronically leak Ca+2. This leakage causes a tonic release of neurotransmitter to the synapses. It is thought that this tonic release is what allows the hair cells to respond so quickly in response to mechanical stimuli. The quickness of the hair cell response may also be due to that fact that it can increase the amount of neurotransmitter release in response to a change as little as 100 μV in membrane potential.
 to ent")
16
Внешние волосковые клетки – умножители звукового сигнала In mammalian outer hair cells, the receptor potential triggers active vibrations of the cell body. This so-called somatic electromotility consists of oscillations of the cells length, which occur at the frequency of the incoming sound and in a stable phase relation. Outer hair cells have evolved only in mammals. They have not improved hearing sensitivity, which reaches similarly exquisite values also in other classes of vertebrates. But they have extended the hearing range from about 11 kHz (maximum in some birds) to about 200 kHz (maximum in some marine mammals). They have also improved frequency selectivity (frequency discrimination), which is of particular benefit for humans, because it enabled sophisticated speech and music. The molecular biology of hair cells has seen considerable progress in recent years, with the identification of the motor protein (prestin) that underlies somatic electromotility in the outer hair cells. Prestin's function is dependent on chloride channel signalling and that it is compromised by the common marine pesticide tributylin (TBT). Because this class of pollutant bioconcentrates up the food chain, the effect is pronounced in top marine predators such as Orcas and toothed whales.

17
Структура улитки (млекопитающие)
")
18
Колебания перегородки улитки

19
Настройка волосковой клетки

20
Нейральные контакты Neurons of the auditory or vestibulocochlear nerve (the VIIIth cranial nerve) innervate cochlear and vestibular hair cells. The neurotransmitter released by hair cells to stimulate the dendrites of afferent neurons is thought to be glutamate. At the presynaptic juncture, there is a distinct presynaptic dense body or ribbon. This dense body is surrounded by synaptic vesicles and is thought to aid in the fast release of neurotransmitter. Nerve fiber innervation is much denser for inner hair cells than for outer hair cells. A single inner hair cell is innervated by numerous nerve fibers, whereas a single nerve fiber innervates many outer hair cells. Inner hair cell nerve fibers are also very heavily myelinated, which is in contrast to the unmyelinated outer hair cell nerve fibers. Efferent projections from the brain to the cochlea also play a role in the perception of sound. Efferent synapses occur on outer hair cells and on afferent (towards the brain) dendrites under inner hair cells. The presynaptic terminal bouton is filled with vesicles containing acetylcholine and a neuropeptide called Calcitonin gene-related peptide (CGRP). The effects of these compounds varies, in some hair cells the acetylcholine hyperpolarized the cell, which reduces the sensitivity of the cochlea locally.
 innervate cochlear and vestibular hair cells. The neurotransmitter released by hair cells to stimulate the dendrites of afferent neurons is thought to b")
21
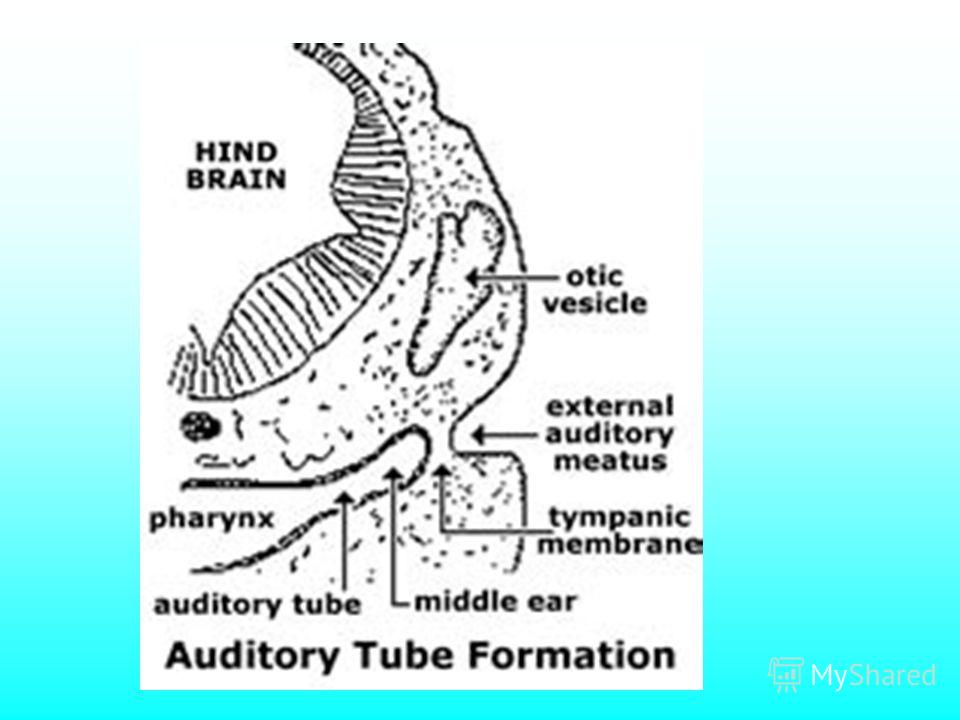
Зародыш млекопитающего

22
Развитие вестибулярного аппарата человека

23
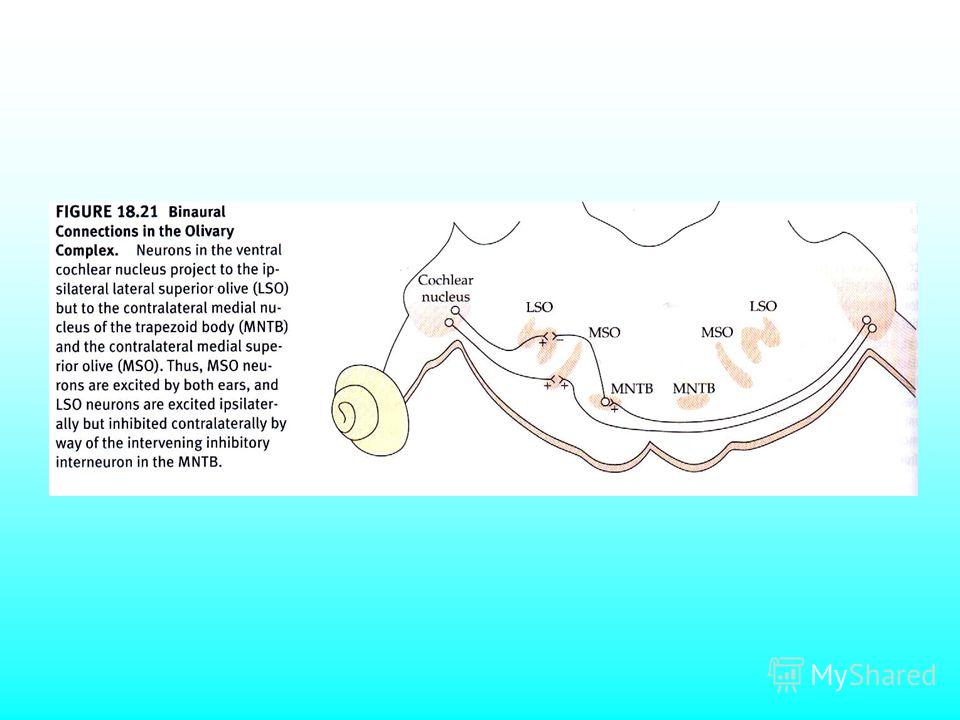
Главные пути передачи звукового сигнала

24
Соматосенсорный кортекс

26
hair cells scar (a) (b) hours before hearing can be damaged loudness range jet takeoff (at 200 ft) rock concert subway, stereo headphones (high volume) motorcycle, lawn mower normal talking quiet background decibels X 1/4 2 8 urban street Громкие звуки могут повреждать волосковые клетки
 (b) hours before hearing can be damaged loudness range jet takeoff (at 200 ft) rock concert subway, stereo headphones (high volume) motorcycle, lawn mower normal talking quiet background 1201008060 decibels 40200 X 1/4 2 8 urban s")
28
ВКУС Вкусовые рецепторы организованы в виде кластеров на языке

29
Вкус Detected by taste buds Taste types –Sour –Salty –Bitter –Sweet –Umami

30
(a) The human tongue (b) Taste bud papillae epithelium of tongue taste receptor cells microvillitaste pore supporting cells nerve fibers to brain
 The human tongue (b) Taste bud papillae epithelium of tongue taste receptor cells microvillitaste pore supporting cells nerve fibers to brain")
31
Клетки вкусовых рецепторов находятся во вкусовых сосочках

32
Развитие вкусовых сосочков

33
Na + Cl – Na + Cl – Na + K+K+ K+K+ K+K+ K+K+ K+K+ K+K+ K+K+ Org – (extracellular fluid) (cytoplasm) axon Нейрон поддерживает ионные градиенты
 (cytoplasm) axon Нейрон поддерживает ионные градиенты")
34
Механизмы передачи вкусовых сигналов

35
Структура вкусовых сосочков. Taste buds (left) are composed of 50–150 taste-receptor cells (TRCs) depending on the species, distributed across different papillae. b, Recent molecular and functional data shown there is no tongue 'map' modalities are present in all areas of the tongue.
 are composed of 50–150 taste-receptor cells (TRCs) depending on the species, distributed across different papillae. b, Recent molecular and functional data shown there is no tongue 'map' modalities are p")
36
Различные модели восприятия вкуса у мышей (Northcutt, 2004)
")
37
a, In the labelled-line model, receptor cells are tuned to respond to single taste modalities sweet, bitter, sour, salty or umami and are innervated by individually tuned nerve fibres. In this case, each taste quality is specified by the activity of non-overlapping cells and fibres. b,c, Two contrasting models of what is known as the 'across- fibre pattern'. This states that either individual TRCs are tuned to multiple taste qualities (indicated by various tones of grey and multicoloured stippled nuclei), and consequently the same afferent fibre carries information for more than one taste modality (b), or that TRCs are still tuned to single taste qualities but the same afferent fibre carries information for more than one taste modality (c). In these two models, the specification of any one taste quality is embedded in a complex pattern of activity across various lines. Recent molecular and functional studies in mice have demonstrated that different TRCs define the different taste modalities, and that activation of a single type of TRC is sufficient to encode taste quality, strongly supporting the labelled-line model.

38
Although many details of the development of the innervation of taste buds are still unknown, it is now clear that taste buds are induced from either ecto- or endodermal epithelia, rather than arising from either placodes or neural crest. At present, there are two developmental models of taste bud induction: The neural induction model claims that peripheral nerve fibers induce taste buds, whereas the early specification model claims that oropharyngeal epithelium is specified by or during gastrulation and that taste buds arise from cell-cell interactions within the specified epithelium. There is now substantial evidence that the early specification model best describes the induction of taste buds.

40
Кора переднего мозга содержит центры обработки первичных сигналов вкуса, слуха и зрения

Еще похожие презентации в нашем архиве:







![Genetics Genetics (from Ancient Greek γενετικός genetikos, genitive and that from γένεσις genesis, origin),[1][2][3] a discipline of biology, is the.](/thumbs/17/1180906/big_thumb.jpg "Genetics Genetics (from Ancient Greek γενετικός genetikos, genitive and that from γένεσις genesis, origin),[1][2][3] a discipline of biology, is the.")


. Centrifugal force (from Latin centrum center and fugere to flee) can generally be any force directed.")
 is an electrical circuit consisting of a resistor, an inductor, and a capacitor, connected in series or in.")



 is the physical theory of measurement in an inertial.")

 trunks in the IP Office.")





