Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Роль «биологических часов» в жизни растения a. The expression of several genes shows circadian rhythms. The genes encoding the light-harvesting chlorophyll-a/b-binding proteins (Lhcb or CAB) and nitrate reductase (NIA2). The clock coordinate the metabolism to maximize photosynthetic yields. b. Cytosolic concentrations of free calcium to oscillate with a circadian rhythm in Arabidopsis c. The clock regulates the phosphorylation of some proteins. Circadian activity of a kinase that phosphorylates phosphoenol pyruvate carboxylase (Kalanchoe fedtschenkoi) d. chloroplast movement (Arabidopsis); e. stomatal opening (Arabidopsis); f. hypocotyl elongation (Arabidopsis): g. cotyledon and leaf movements (Arabidopsis): h. petal opening (Kalanchoe): i. synchronizing developmental processes such as flowering time. Mutations in all the putative clock-associated genes cause altered photoperiodic control of flowering..
 and nitrate reductase (NIA2). The clock coordinate the meta")
2
Компоненты системы «биологических часов» Components of a circadian system. This simple model includes an input pathway (from light and/or temperature) to a circadian oscillator. The oscillator generates signals that are transduced via output pathways to produce overt circadian rhythms (output). The output is depicted as two idealized rhythms (red and green lines) with different phases. Yellow and grey boxes represent light and dark (diurnal) intervals, respectively. Under diurnal conditions, the period of the oscillator (the time between comparable points in the repeating cycle) matches the period of the entraining cycles. Under constant onditions, the clock oscillates with an ndogenous period close to 24 h. Amplitude is half the distance between the peak and trough.
 to a circadian oscillator. The oscillator generates signals that are transduced via output pathways")
3
Молекулярная модель эукариотического циркадного осциллятора a. General scheme showing the core feedback loop central to eukaryotic circadian oscillators. Paired positive elements (CLOCK and CYC in Drosophila, WC-1 and WC-2 in Neurospora, CLOCK and BMAL in mice) form heterodimers that act as transcriptional activators to induce clock gene expression. The protein products (negative elements) of the clock genes (PER and TIM in Drosophila, FRQ in Neurospora, mCRYs and mPERs in mice) in turn block the action of the positive elements, thus indirectly repressing their own expression. b. Мodel of the Neurospora circadian system. The WC-1–WC-2 heterodimer activates expression of the clock gene FRQ. The FRQ protein has two roles. In one loop, it acts as a negative regulator of the WC-1–WC-2 heterodimer. In a second loop, FRQ acts as a positive regulator of WC-1 protein synthesis from existing WC-1 mRNA. Positive elements are depicted in blue and negative elements in red. Because FRQ acts as both a positive and negative element, it is depicted in red and blue. a. General scheme showing the core feedback loop central to eukaryotic circadian oscillators. Paired positive elements (CLOCK and CYC in Drosophila, WC-1 and WC-2 in Neurospora, CLOCK and BMAL in mice) form heterodimers that act as transcriptional activators to induce clock gene expression. The protein products (negative elements) of the clock genes (PER and TIM in Drosophila, FRQ in Neurospora, mCRYs and mPERs in mice) in turn block the action of the positive elements, thus indirectly repressing their own expression. b. Мodel of the Neurospora circadian system. The WC-1–WC-2 heterodimer activates expression of the clock gene FRQ. The FRQ protein has two roles. In one loop, it acts as a negative regulator of the WC-1–WC-2 heterodimer. In a second loop, FRQ acts as a positive regulator of WC-1 protein synthesis from existing WC-1 mRNA. Positive elements are depicted in blue and negative elements in red. Because FRQ acts as both a positive and negative element, it is depicted in red and blue.

4
Модель циркадной системы Arabidopsis Тhe oscillator and the input and output pathways are fuzzy. Proteins have been placed within the shaded area according to how close they might be to the oscillator. The darker the shading (towards the center), the closer to the oscillator. Thus, CCA1, LHY and TOC1, which might be part of the oscillator itself are located at the center of the shaded area. Similarly, ZTL and FKF1, although assigned to the input pathways, are also placed near the oscillator because they might interact with phosphorylated oscillator proteins, targeting them for degradation. Oscillator proteins might be phosphorylated by CK2. The PIF3–PHYB complex might also interact directly with the CCA1 promoter and is therefore placed close to the oscillator. The green arrows indicate that components such as PHYB and GI, assigned as inputs or as part of input pathways, respectively, are themselves affected by the clock. Many of the putative clock components show circadian rhythmicity of transcript levels but the available evidence suggests that ZTL transcripts do not cycle.
, the")
5
Мутанты по индукции флорального морфогенеза LFY

6
«Круговое» строение цветка

7
Мутанты по структуре цветка сходны у разных растений

8
АВС-схема генетики развития цветка (теория «войны позиций)
")
9
Мутанты арабидопсиса с точки зрения АВС-системы

10
Схема строения мутантов арабидопсиса по структуре цветка

11
И еще раз АВС-схема мутантов – но в цветном варианте

12
Предполагаемая схема эволюции АВС-системы у растений

13
Сильно упрощенная схема иерархии генов флорального морфогенеза Гены с MADS-боксом изображены в квадратиках. AG - AGAMOUS; AG - AGAMOUS; AGL - AGAMOUS-LIKE; AGL - AGAMOUS-LIKE; AP – APETALA AP – APETALA BEL1 – BELL BEL1 – BELL CAL - CAULIFLOWER; CAL - CAULIFLOWER; CO - CONSTANS; CO - CONSTANS; ELF1 - EARLY FLOWERING1; ELF1 - EARLY FLOWERING1; LD - LUMINIDEPENDENS; LD - LUMINIDEPENDENS; LFY - LEAFY; LFY - LEAFY; LUG - LEUNIG; LUG - LEUNIG; NAP - NAC-LIKE, NAP - NAC-LIKE, PI - PISTILLATA; PI - PISTILLATA; SIN1 - SHORT INTEGUMENTS1; SIN1 - SHORT INTEGUMENTS1; SUP - SUPERMAN; SUP - SUPERMAN; UFO - UNUSUAL FLORAL ORGANS; UFO - UNUSUAL FLORAL ORGANS; TFL - TERMINAL FLOWER. TFL - TERMINAL FLOWER. Гены с MADS-боксом изображены в квадратиках. AG - AGAMOUS; AG - AGAMOUS; AGL - AGAMOUS-LIKE; AGL - AGAMOUS-LIKE; AP – APETALA AP – APETALA BEL1 – BELL BEL1 – BELL CAL - CAULIFLOWER; CAL - CAULIFLOWER; CO - CONSTANS; CO - CONSTANS; ELF1 - EARLY FLOWERING1; ELF1 - EARLY FLOWERING1; LD - LUMINIDEPENDENS; LD - LUMINIDEPENDENS; LFY - LEAFY; LFY - LEAFY; LUG - LEUNIG; LUG - LEUNIG; NAP - NAC-LIKE, NAP - NAC-LIKE, PI - PISTILLATA; PI - PISTILLATA; SIN1 - SHORT INTEGUMENTS1; SIN1 - SHORT INTEGUMENTS1; SUP - SUPERMAN; SUP - SUPERMAN; UFO - UNUSUAL FLORAL ORGANS; UFO - UNUSUAL FLORAL ORGANS; TFL - TERMINAL FLOWER. TFL - TERMINAL FLOWER.

14
То же, но чуть подробнее

15
Патогенов много и они специфичны для разных органов растения Проникновение гриба после прорастания спор через поверхность листа ячменя Проникновение гриба Rhynchosporium secalis после прорастания спор через поверхность листа ячменя

16
Примеры защитных соединений «первого круга» обороны: алкалоиды

17
Примеры защитных соединений «первого круга» обороны: изопреноиды

18
Еще примеры защитных соединений «первого круга» обороны

19
Примеры защитных соединений «второго круга» обороны: цианогенные гликозиды, глюкозинолаты.

20
Этапы ответа растения на вторжение патогена

21
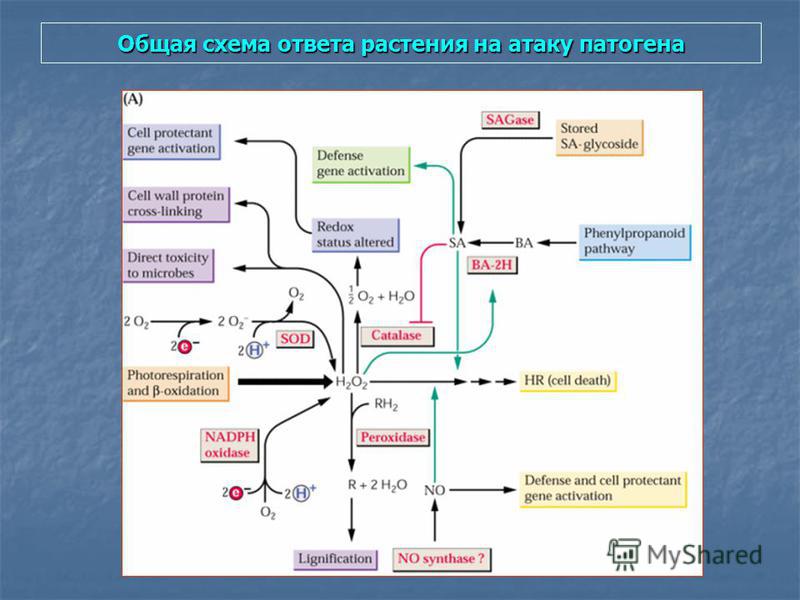
Общая схема ответа растения на атаку патогена

23
Взаимодействие патогена и растения

24
Протеин-киназы, участвующие в трансдукции защитного сигнала Киназы, обнаружены в: табаке (красный), томатах (оранжевый), люцерне (синий), петрушке (желтый), рисе (коричневый), Arabidopsis (зеленый).
, томатах (оранжевый), люцерне (синий), петрушке (желтый), рисе (коричневый), Arabidopsis (зеленый).")
25
Общая схема ответа растения на патогенез Общая схема ответа растения на патогенез

26
Возникновение системной устойчивости растений

27
Cхема рецепции и передачи сигналов при патогенезе

Еще похожие презентации в нашем архиве:
, на С-конце.")


. Centrifugal force (from Latin centrum center and fugere to flee) can generally be any force directed.")


. Like a set, it contains members (also called elements, or terms),")

. Family How could you describe the word family? First of all family means a close unit of parents and their.")





 objects.")





