Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Вторичная структура белков PEPTOCOCCUS AEROGENES FERREDOXIN HORMONE GLUCAGON HYDROLASE (SERINE PROTEINASE )
")
2
Первичная структура может свернуться в пространстве и принять ту или иную пространственную структуру, которая называется вторичной структурой. Вторичная структура (1)Возможности относительно свободного вращения вокруг некоторых связей (2) Возможности взаимодействий между близкими по цепи, но ковалентно несвязанными атомами
Возможности относительно свободного вращения вокруг некоторых связей (2) Возможности")
3
. Образование водородных связей (показаны стрелками) В принципе, возможны такие спирали, стянутые Н-связями: 2 7, 3 10, 4 13 (обычно именуемая ), 5 16 (или ). В названии "2 7 " - "2" означает связь со 2-м по цепи остатком, а "7" - число атомов в цикле (O......H-N-C'-C -N-C'), замыкаемом этой связью.
 В принципе, возможны такие спирали, стянутые Н-связями: 2 7, 3 10, 4 13 (обычно именуемая ), 5 16 (или ). В названии")
4
Правые (R) спирали Левые (L) спирали Показан отсчет положительного угла: близкая к нам стрелка вращается против хода часов
 спирали Левые (L) спирали Показан отсчет положительного угла: близкая к нам стрелка вращается против хода часов")
5
(б) Схематическое изображение одного витка -спирали, вид с торца. Стрелка показывает поворот спирали (в расчете на один остаток) по мере ее приближения. а) Атомарная структура. R - боковые группы. Голубые линии - водородные связи Правая -спираль
 Схематическое изображение одного витка -спирали, вид с торца. Стрелка показывает поворот спирали (в расчете на один остаток) по мере ее приближения. а) Атомарная структура. R - боковые группы. Голубые линии - водородные связи Правая -спираль")
6
Карта разрешенных и запрещенных значений углов для всех гипотетически возможных спиралей полипептидной цепи: 2 7, 3 10, 4 13 ( ), 5 16 ( ), правые (R) и левые (L).
, 5 16 ( ), правые (R) и левые (L).")
7
Модель -спиральной конформации полипептидной цепи. -спираль -спираль Водородные связи формируются «параллельно» направлению полипептидного остова вдоль оси спирали!

8
Параметры -спирали: 1. 3,6 аминокислотных остатков на виток спирали 2.Шаг спирали Р = 5.4Å 3. Трансляция на один виток d = 1,5Å 4.Радиус спирали r = 2,3Å 5. Водородные связи образуются между 1 и 4 пептидными группами

9
6. = -57 о, = -47 о Параметры -спирали

10
-структура -структура Водородных связей в -структуре – «перпендикулярно» направлению полипептидного остова. Антипараллельная Параллельная

11
-структуры: Характеристики -структуры: 6 полипептидных цепей 6 аминокислотных остатков в цепи 6 полипептидных цепей 6 аминокислотных остатков в цепи

12
t d Длина листа (6 остатков) d =20 Å Ширина листа (6 цепей) t =25Å Характеристики -структуры: -структуры:
 d =20 Å Ширина листа (6 цепей) t =25Å Характеристики -структуры: -структуры:")
13
-структуры: Характеристики -структуры: Антипараллельные = -139 о = -139 о, = +135 о параллельные = -119 о = -119 о, = +113 о

14
-лист складчатый -лист скрученный -листы скручены из-за того, что скручены отдельные -тяжи Скрученность отдельного -тяжа - левая (у L аминокислот), боковые группы тяжа поворачиваются по часовой стрелке (на -165 o на каждый остаток) по мере приближения тяжа. Боковые группы расположены на складках; каждая обращена в ту же, что и складка, сторону, т.е. направленные вниз и вверх боковые группы чередуются вдоль -тяжа.
, боковые группы тяжа поворачиваются по часовой стрелке (на -165 o на каждый остаток) по мере приближени")
15
Конформация идеальной (не скрученной) параллельной и антипараллельной -структуры для поли(Gly), и усредненная конформация реальной (сложенной из L аминокислот) скрученной (twist) -структуры. Скрученность -листа Пунктир показывает область энергетического минимума для отдельно взятого остатка Ala; контуром показаны границы области разрешенных конформаций этого остатка. Диагональ -карты соответствует плоской периодичной структуре, имеющей 2 остатка на виток (n=2). Над диагональю лежат левые (L) спирали, под ней - правые (R).
 параллельной и антипараллельной -структуры для поли(Gly), и усредненная конформация реальной (сложенной из L аминокислот) скрученной (twist) -структуры. Скрученность -листа Пунктир показывает область энергетическ")
16
- изгибы - изгибы Конформация -изгибов типа I (а) и II (б)
 и II (б)")
17
- изгибы - изгибы Конформация -изгибов типа I (а) и типа II (б)
 и типа II (б)")
18
Третий тип спирали – пролиновые спирали: полипролин I и полипролин II. Пролиновые спирали

19
Первая – полипролин I - левая спираль, содержит 10 остатка на 3 витка; все пептидные связи в цис- конформации. Вторая – полипролин II - левая спираль, содержит 3 остатка на виток; пептидные связи в транс-положении. Спираль полипролин II имеет более вытянутую цепь, чем -спираль: смещение вдоль оси спирали 3.12 Å на остаток по сравнению с 1.5 Å для -спирали. В нормальных физиологических условиях спираль полипролин II более стабильна.

20
Основные геометрические параметры наиболее распространенных в белках вторичных структур Структура H-связьОстаток/ виток Смещение/ остаток (Å) Спираль R (4 13 ) R CO 0 HN Спираль (3 10 ) R CO 0 HN Лист -паралл. меж цепей * Лист - анти меж цепей * Спираль Poly(Pro) IIнет
 Спираль R (4 13 ) R CO 0 HN +4 +3.61.5-57 0 -47 0 Спираль (3 10 ) R CO 0 HN +3 +32.0-50 0 -25 0 Лист -паралл")
21
для пролина для аланина для больших остатков P Карты Рамачандрана для наиболее распространенных в белках вторичных структур

22
Экспериментальное определение вторичной структуры белков Спираль ввиду ее симметрии легко распознать и описать. На карте электронной плотности, полученной методом РСА, спирали видны как светлые круги (вид с торца) или как периодически расположенные светлые пятна (вид спирали сбоку). Метод РСА (ренгено- структурный анализ)
 или как периодически расположенные с")
23
Сближенность ядер атомов Н, наиболее характерная для ЯМР ядерный магнитный резонанс антипараллельной -структуры параллельной -структуры -спирали

24
Экспериментальное определение вторичной структуры белков Для полилизина в форме -спирали (a), -структуры (b) и неупорядоченного клубка (r). Спектры КД ИК-спектры пропускания в тяжелой воде (D 2 O)
, -структуры (b) и неупорядоченного клубка (r). Спектры КД ИК-спектры пропускания в тяжелой воде (D 2 O)")
25
Стабильность вторичной структуры белков и кинетика ее образования

26
α-спирали следующая водородная связь, (СО) 1 …(HN) 5, дополнительно фиксирует конформацию еще одного остатка - остатка 4 ; связь (СО) 2 … (HN) 6 дополнительно фиксирует остаток 5, и т.д. Таким образом, если в спирали фиксировано n остатков, то их фиксирует n-2 водородные связи Первая водородная связь в α-спирали, (СО) 0 …(HN) 4, фиксирует конформации трех остатков - 1, 2, 3;
 1 …(HN) 5, дополнительно фиксирует конформацию еще одного остатка - остатка 4 ; связь (СО) 2 … (HN) 6 дополнительно фиксирует остаток 5, и т.д. Таким образом, если в спирали фиксировано n остатков, то их фик")
27
F = F F клуб. = (n-2)f H nTS = -2f H + n(f H TS ) Свободная энергия образования α-спирали из клубка в водном окружении f H - свободная энергия водородной связи в α-спирали; и энергия, и энтропия перестроек водородных связей в водном окружении, S - потеря энтропии при фиксации одного остатка в спирали. В F есть два члена. (1) -2f H - не зависит от длины спирали; f INIT = -2f H - свободная энергия инициации спирали; f INIT - суммарная свободная энергия обеих границ спирали с клубком учитывает и инициацию, и терминацию спирали. (2 ) n(f H - TS ) - прямо пропорционален длине спирали. f EL = (f H - TS ) - свободная энергия элонгации спирали на 1 остаток. В общем виде, F = f INIT + n. f EL
f H nTS = -2f H + n(f H TS ) Свободная энергия образования α-спирали из клубка в водном окружении f H - свободная энергия водородной связи в α-спирали; и энергия, и энтропия перестроек водородных связей в водном окружении, S - по")
28
Отношение вероятности спирального состояния цепи из n остатков к ее клубковому состоянию, равно exp(- F /kT) = exp(-f INIT /kT). [exp(-f EL /kT)] n = s n s = exp(-f EL /kT) - фактор элонгации спирали: s = exp(-f EL /kT); = exp(-f INIT /kT) - фактор инициации спирали: = exp(-f INIT /kT);
![Отношение вероятности спирального состояния цепи из n остатков к ее клубковому состоянию, равно exp(- F /kT) = exp(-f INIT /kT). [exp(-f EL /kT)] n = s n s = exp(-f EL /kT) - фактор элонгации спирали: s = exp(-f EL /kT); = exp(-f INIT /kT) - фактор и](http://images.myshared.ru/5/330382/slide_28.jpg "Отношение вероятности спирального состояния цепи из n остатков к ее клубковому состоянию, равно exp(- F /kT) = exp(-f INIT /kT). [exp(-f EL /kT)] n = s n s = exp(-f EL /kT) - фактор элонгации спирали: s = exp(-f EL /kT); = exp(-f INIT /kT) - фактор и")
29
Как образуется спираль при изменении условий среды (температуры, растворителя и т.д.): переходом "все-или-ничего" или постепенно?
: переходом все-или-ничего или постепенно?")
30
Теорема Ландау: В системе, где обе фазы одномерны, фазовый переход первого рода невозможен «Одномерность» - это значит, что размер границы, "стыка" фаз не зависит от размера фрагментов этих фаз. В этом смысле и спираль, и клубок в полимере одномерны.

31
«Происходит как фазовый переход первого рода". Это значит, что при температуре перехода стабильной может быть либо одна, либо другая фаза, но перемешивание фаз (например, льда и воды) ведет к повышению свободной энергии, и потому нестабильно.
 ведет к повышению свободной энергии, и потому нестабильно.")
32
Выгодно ли сосуществование фаз в трехмерной системе? Нет. Почему? Рассмотрим температуру, где безграничная вода и безграничный лед имеют равную свободную энергию – это условие "точки перехода". Если в воде плавает льдина из n молекул, свободная энергия границы пропорциональна n 2/3, где n 2/3 - характерное число пограничных молекул, > 0 - энергия границы в расчете на одну из пограничных молекул. Если < 0, "перемешивание" (оно в этом случае термодинамически выгодно) произойдет на молекулярных масштабах, и двух фаз вообще не будет.
0 - энергия границы в расчете на одну из пограничных молекул. Если < 0, "перемешивание" (оно в этом случае термодинамически выгодно) произойдет на молекулярных масштабах, и двух фаз вообще не будет.">

33
Поверхность льдинки повышает свободную энергию на n 2/3. Позиционная энтропия льдинки ( она может находиться в разных точках сосуда ) не превосходит величину порядка k. ln(N), если в сосуде N молекул - N точек, с которых может начинаться льдинка. F [ n 2/3 - kТ. ln(N)] (cвободная энергия льдинки) Логарифм медленно растет для больших N. Если льдинка занимает заметную часть сосуда ( n ~ N/10), то ln(N) (23) мал по сравнению с (N/10) 2/3, то есть в F доминирует граничный член n 2/3, а он противится ее образованию. В трехмерной системе макроскопические фазы разделяются, и фазовый переход первого рода в ней возможен
 не превосходит величину порядка k. ln(N), если в сосуде N молекул - N точек, с которых может начинаться льдинка. F [")
34
Выгодно ли сосуществование фаз в одномерной системе? Оказывается, да. Рассмотрим температуру "середины перехода", где спираль и клубок имеют равную свободную энергию, т.е. f EL =0. Свободная энергия обеих границ спирали и клубка, f INIT, не зависит ни от размера спирали, ни от размера клубка. Позиционная энтропия спирали длины n в цепи длины N равна S = k. ln(N-n) и свободная энергия этой спирали F hel f INIT - kТ. ln(N-n)

35
При больших N член с ln(N-n) всегда доминирует в выражении F hel = f INIT - kТ. ln(N-n) даже если n ~ 0.9 N Логарифмический член ln(N-n) понижает свободную энергию и способствует внедрению спирали в клубок (и, точно так же, - клубка в спираль). Поэтому в одномерной системе фазы не разделяются, они стремятся перемешаться и фазовый переход первого рода, или типа "все-или-ничего", невозможен при достаточно большой длине цепи. Теорема Ландау доказана.
 всегда доминирует в выражении F hel = f INIT - kТ. ln(N-n) даже если n ~ 0.9 N Логарифмический член ln(N-n) понижает свободную энергию и способствует внедрению спирали в клубок (и, точно так же, - клубка в спираль). Поэто")
36
Для большинства аминокислот f INIT 4 ккал/моль, и 0.001, n 0 ~ 30. Для измерения влияния отдельных аминокислотных остатков на стабильность спиралей используют короткие (длиной n 0 ~ 30 или менее) полипептиды. В них может образоваться только одна спираль, и оценить влияние каждой аминокислотной замены на спиральность здесь наиболее просто.
 полипептиды. В них может образоваться только одна спир")
37
Вклад аминокислотного остатка в стабильность спирали лежит в следующих пределах: аланин - самый "спиралеобразующий" остаток: f EL -0.4 ккал/моль, т.е. s 2; глицин - самый "спиралеразрушающий" остаток: f EL +1 ккал/моль, т.е. s 0.2; пролин - иминокислота (не имеющей NH группы, которая должна завязывать структурообразующую водородную связь в α-спирали) величина s , но точно не измерена.

38
Это пример не фазового, а кооперативного перехода: при малой величине фактора инициации спирали (

39
Рассмотрим скорость образования вторичной структуры в пептидах Такой барьер похож на активационный барьер в химических реакциях, только в этом случае он имеет и энергетическую, и энтропийную составляющие. "Медленность" процессов связана с преодолением высокого свободно-энергетического барьера. Особенно это типично для переходов типа "все или ничего", где свободно-энергетический барьер разделяет две фазы и он всегда значителен. Преодоление свободно-энергетического (активационного) барьера # при переходе из состояния "А" в состояние "В". F # - свободная энергия барьера - перехoдного состояния. Кинетика

40
α-спирали образуются быстро: за наносекунды (10 -9 с) α-спираль охватывает пептид из остатков, т.е. спирали нарастают со скоростью порядка 1 остаток за несколько наносекунд. Скорость образования α-спирали зависит от скорости, с которой она нарастает, и от скорости, с которой в цепях появляются первые зародыши спиральной структуры. Инициация α-спирали требует преодоления активационного барьера, поэтому для образования первого витка спирали необходимо больше времени, чем ее последующее удлинение еще на 1 виток. Т.е. элонгация α-спирали идет быстро, а все наблюдаемое время тратится на ее инициацию.
 α-спираль охватывает пептид из 20-30 остатков, т.е. спирали нарастают со скоростью порядка 1 остаток за несколько наносекунд. Скорость образования α-спирали зависит от скорости, с которой она нара")
41
Характерная зависимость свободной энергии спирали ( F a ) от числа вовлеченных в нее аминокислотных остатков (N) при разных свободных энергиях ее элонгации (f EL ). При f EL 0 - спираль любой длины нестабильна и, следовательно, не образуется.
 от числа вовлеченных в нее аминокислотных остатков (N) при разных свободных энергиях ее элонгации (f EL ). При f EL 0 - спираль любой длины нестабильна и, следовательно, не образуется.")
42
Кинетика образования α-спиралей относительно проста: они всегда образуются быстро. Кинетика образования -структуры много сложнее и интереснее. -структура в полипептидах часто образуется очень медленно – и за часы, и за недели (но и за миллисекунду). Почему? Белки с -структурой сворачиваются не медленнее α- спиральных белков. Как они успевают? И что отвечает за аномалии в образовании -структуры в полипептидах: медленная инициация или медленная элонгация? "Аномальная" (по сравнению с переходом спираль- клубок) кинетика образования -структуры связана с тем, что она - двумерный объект, а не одномерный, как спираль или клубок.
.")
43
Схема -листа. Аминокислотные остатки внутренних - участков - черные кружки, краевых -участков - пустые кружки; изгибы (или петли), соединяющие -участки, - уголки. Сценарий роста -листа. Самая нестабильная структура помечена #. Край -листа сложен из краевых -тяжей и изгибов (петель), соединяющих все -тяжи. б
, соединяющие -участки, - уголки. Сценарий роста -листа. Самая нестабильная структура помечена #. Край -листа сложен из к")
44
Находящиеся на краю листа звенья цепи имеют меньше контактов с другими, чем внутренние звенья листа. Т.е., край -листа (как и граница другой фазы - капли воды, льдинки или α-спирали) имеет повышенную свободную энергию. В отличие от α-спирали (и в сходстве с каплей) -лист не одномерен, размер его края (и его свободная энергия) растет с числом вовлеченных в лист звеньев цепи. Поэтому переход клубок - -структура фазовый переход первого рода, как образование капли или льдинки. В результате на пути образования -листа возникает высокий (особенно при образовании лишь слегка стабильной -структуры) свободно-энергетический барьер, который во много раз замедляет инициацию его сворачивания.
 имеет повышенную свободную энергию. В отличие от α-спирали (и в схо")
45
Пусть свободная энергия клубка принята за 0 (начало отсчета), тогда f - свободная энергия остатка в центре -листа, f + f - свободная энергия остатка на его краю ( f - краевой эффект), U - свободная энергия изгиба. Если -лист образуется, то он стабилен ( f 0 и U > 0 ). Край -листа сложен из (а) краевых -тяжей, и (б) изгибов или петель, соединяющих все -тяжи. а б б б а
, тогда f - свободная энергия остатка в центре -листа, f + f - свободная энергия остатка на его краю ( f - краевой эффект), U - свободная энергия изгиба. Если -лист образуется, то он стабиле")
46
В кинетике образования -структуры надо различать два случая: 1) f + f
 f + f")
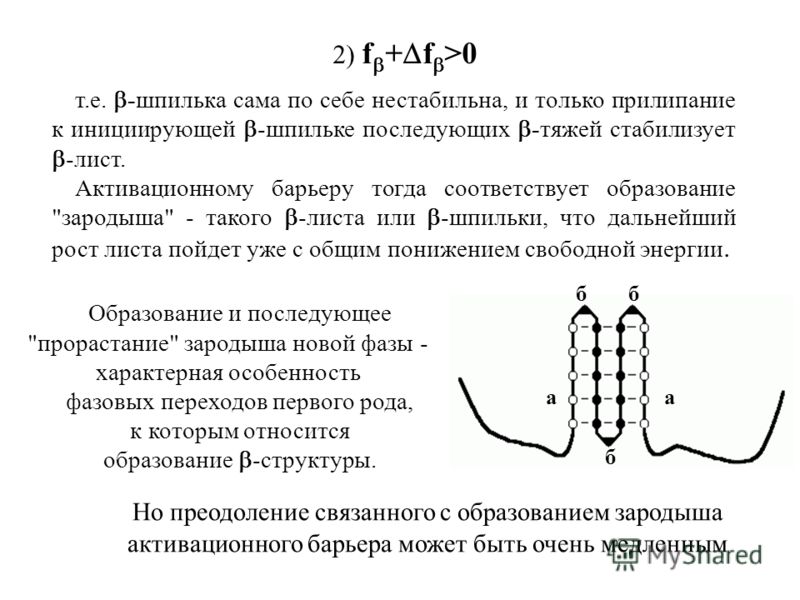
0 т.е. -шпилька сама по себе нестабильна, и только прилипание к инициирующей -" title="Образование и последующее "прорастание" зародыша новой фазы - характерная особенность фазовых переходов первого рода, к которым относится образование -структуры. 2) f + f >0 т.е. -шпилька сама по себе нестабильна, и только прилипание к инициирующей -" class="link_thumb">
47
Образование и последующее "прорастание" зародыша новой фазы - характерная особенность фазовых переходов первого рода, к которым относится образование -структуры. 2) f + f >0 т.е. -шпилька сама по себе нестабильна, и только прилипание к инициирующей -шпильке последующих -тяжей стабилизует -лист. Активационному барьеру тогда соответствует образование "зародыша" - такого -листа или -шпильки, что дальнейший рост листа пойдет уже с общим понижением свободной энергии. а б б б а Но преодоление связанного с образованием зародыша активационного барьера может быть очень медленным
0 т.е. -шпилька сама по себе нестабильна, и только прилипание к инициирующей -">
0 т.е. -шпилька сама по себе нестабильна, и только прилипание к инициирующей -шпильке последующих -тяжей стабилизует -лист. Активационному барьеру тогда соответствует образование "зародыша" - такого -листа или -шпильки, что дальнейший рост листа пойдет уже с общим понижением свободной энергии. а б б б а Но преодоление связанного с образованием зародыша активационного барьера может быть очень медленным">
0 т.е. -шпилька сама по себе нестабильна, и только прилипание к инициирующей -" title="Образование и последующее "прорастание" зародыша новой фазы - характерная особенность фазовых переходов первого рода, к которым относится образование -структуры. 2) f + f >0 т.е. -шпилька сама по себе нестабильна, и только прилипание к инициирующей -">
48
Рассмотрим простейший сценарий роста стабильного -листа: образование инициирующей -шпильки из изгиба и двух -тяжей, по N звеньев (аминокислот) в каждом; образование последующего изгиба на ее конце; прилипание к листу -тяжа из N звеньев; образование последующего изгиба; прилипание -тяжа; и т. д. Цикл "прилипание нового -тяжа и образование нового -изгиба" изменяет суммарную свободную энергию на Nf +U. В результате такого цикла свободная энергия должна снижаться (тогда сворачивание цепи идет быстро), и прилипающий -тяж (и тот, к которому он прилипает) должен состоять не менее чем из N min = U/(-f ) звеньев.
 в каждом; образование последующего изгиба на ее конце; прилипание к листу -тяжа из N звеньев; образование п")
49
«Переходным» - самым нестабильным состоянием при образовании -структуры - это -шпилька с последующим изгибом (#). Так как стабильность шпильки падает с удлинением, а -тяж в инициирующей шпильке должен состоять как минимум из N min звеньев N min = U/(-f ), то F min инициирующей -шпильки равна F # = U + 2N min (f + f ) + U = 2(U f )/(- f ). Это свободная энергия переходного состояния при образовании -листа по нашему сценарию. f - свободная энергия остатка в центре -листа, U - свободная энергия изгиба. f + f - свободная энергия остатка на его краю ( f - краевой эффект),
. Так как стабильность шпильки падает с удлинением, а -тяж в инициирующей шпильке должен состоять как минимум из N min звеньев N min = U/(")
50
Как меняется минимальная свободная энергия -листа с его увеличением? Пока лист мал, доминируют краевые эффекты, и свободная энергия повышается. В большом листе доминируют внутренние остатки, и его свободная энергия падает. Кривые относятся к разным свободным энергиям f внутренних звеньев - листа. # -точка максимума величины F opt по ходу роста - листа. При росте - листа (в отличие от роста - спирали) максимум его свободной энергии не приходится на самое начало этого процесса. Общий ход изменения минимальной свободной энергии - листа F opt с увеличением числа N звеньев в нем.

51
При малых -f время образования структуры огромно, в пределе - бесконечно велико. Общий вид зависимости характерного времени образования -листа от стабильности -структуры Так как, согласно теории переходного состояния, время процесса t зависит от свободной энергии переходного (самого нестабильного промежуточного) состояния F # как t ~. exp(+F # /kT), где - время элементарного шага (элонгации -листа на один остаток; скорость элонгации -листа качественно не отличается от скорости элонгации спирали), тогда характерное время образования листа при малых -f есть t ~. exp[A/(-f )], где A - некая константа.

52
Итак: малостабильная -структура должна образовываться очень медленно, и причина не в медленной элонгации, а в медленной инициации: трудно собрать свободную энергию из теплового движения и преодолеть высокий барьер. Так объясняется и экспериментально наблюдаемая, очень медленная скорость образования -структуры в неагрегирующих полипептидах, и резкий ее рост с повышением стабильности -структуры. В то же время стабильные -листы и шпильки (они и наблюдаются в белках) должны образовываться довольно быстро, примерно как -спираль.

53
Крайне медленная инициация - общее свойство фазовых переходов первого рода, когда возникающая фаза находится на грани стабильности. Это связано с большой свободной энергией обширной границы раздела фаз. -структура образуется фазовым переходом первого рода со всеми вытекающими последствиями. Наоборот, -спираль не образуется фазовым переходом первого рода: граница спирали, в отличие от границы -структуры (или льдинки), не растет с ее размером. Поэтому барьер, который нужно преодолеть при сворачивании спирали, всегда имеет конечную, небольшую величину, и она может успеть "проинициироваться" за микросекунду.

54
Вопрос: при каких характерных длинах цепи начинается смешение клубковой и спиральной фаз? Рассмотрим цепь из N звеньев при температуре "середины перехода", где спираль и клубок имеют равную свободную энергию: f EL = 0. Свободная энергия инициации спирали - f INIT. Число возможных положений спирали в цепи из N звеньев порядка N 2 /2 (спираль может начинаться и кончаться в любом месте, ее длина не менее трех остатков). Ни расположение спирали в цепи, ни ее длина при f EL =0 не влияют на свободную энергию спирали.

55
Смешение клубковой и спиральной фаз начинается в фрагментах длины N ~ n 0, и величина n 0 получается из уравнения f INIT - 2kТ. ln(n 0 ) = 0 Полная свободная энергия внедрения фрагмента новой фазы - спирали - в цепь длины N : f INIT - 2kТ. ln(N). и характерная длина кусков спирали и клубка в середине перехода n 0 = exp(+f INIT /2kТ) = - 1/2
 = 0 Полная свободная энергия внедрения фрагмента новой фазы - спирали - в цепь длины N : f INIT - 2kТ. ln(N). и х")
Еще похожие презентации в нашем архиве:
 А.Б.Рахманинова, Д.А.Равчеев, 2009.")



















